NB 2-4 details
videos
NB 2-4 delaminates as an S4 NB.
No information about the lineage derived from NB 2-4 is available from other insects.
NB 2-4 is currently known to express 5 molecular markers at its birth: huckebein (hkb) (Doe, 1992; Chu-LaGraff et al, 1995); muscle-specific homeobox gene 1 (msh) (Buescher et al, 1996; Isshiki et al, 1997); mirror-lacZ (mrr-lacZ) (Broadus et al, 1995; McNeill et al, 1997); eagle (egl) (Higashijima et al, 1996); pox-neuro (poxn) ( Bopp et al, 1989). One cell cycle later (at S5) it adds seven-up-lacZ (svp-lacZ) expression (Broadus et al, 1995), Klumpfuss (Klu) expression (Yang et al, 1997), and castor (cas) expression (Cui and Doe, 1992, 1995).
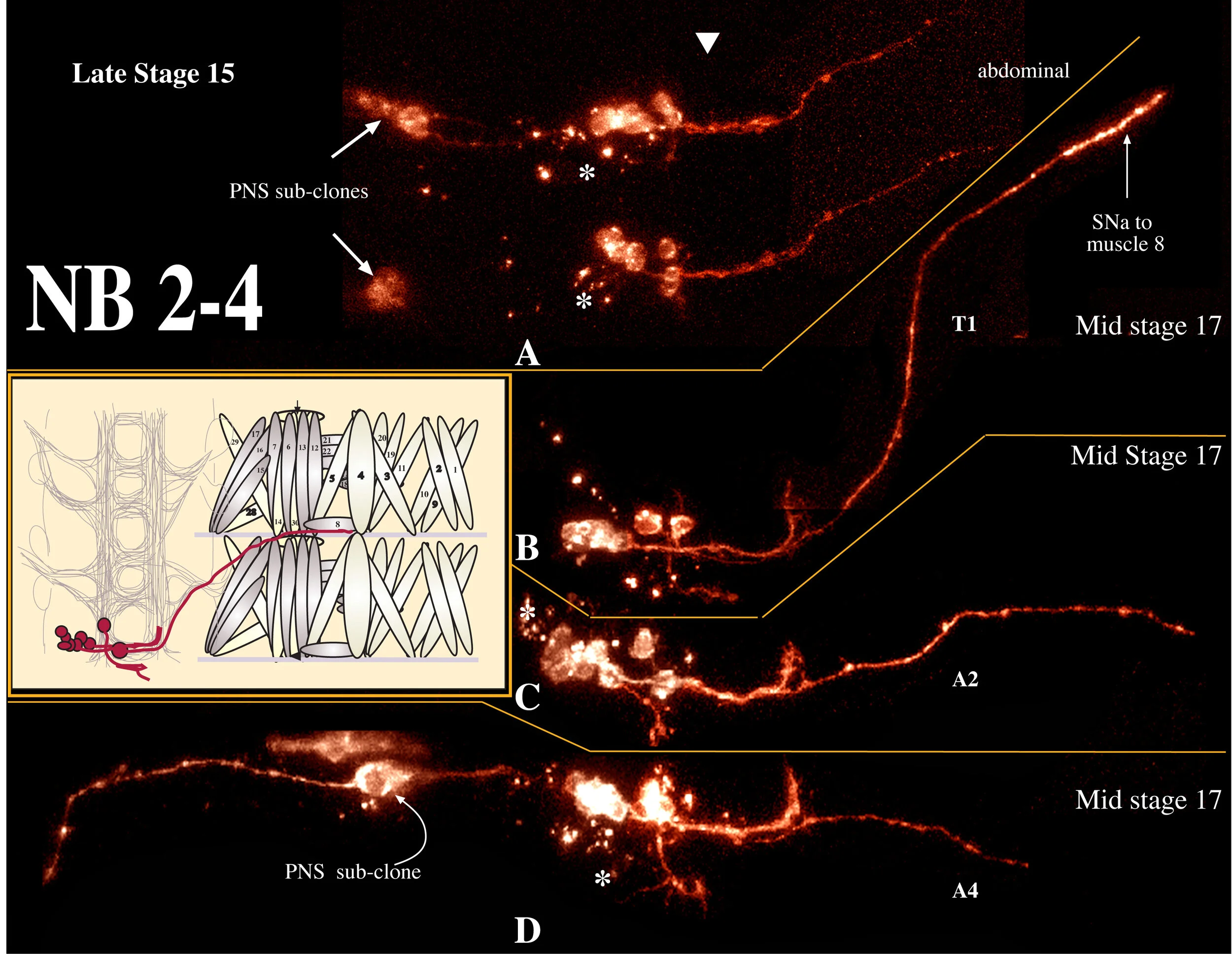
NB 2-4 has been described by Schmidt et al. (1997) as producing 2 motoneurons that project contralaterally out the anterior root of the ISN, 7-8 interneurons with ipsilateral posterior projections, and an obligate epidermal subclone. We generated 8 clones from NB 2-4, and scored 3 at stage 16 (1 thoracic, 2 abdominal; averaging 14 cells per clone) and 5 at stage 17 (2 thoracic, 3 abdominal; averaging 10 cells per clone). As expected from the decline in cell numbers, we detected significant amounts of cell death in this clone (Fig. 2-4, asterisks). We detected one motoneuron and a cluster of local interneurons; 2/8 clones had epidermal cells and 3/8 clones had a PNS subclone.
A. Motoneurons:
Schmidt et al, (1997) report that 2 cells migrate medially in abdominal clones (but not in thoracic clones), that these two cells are motoneurons exiting the CNS via the ISN anterior root, and that the contralaterally projecting fibers from this clone are derived from these two motoneurons. We find only one motoneuron to be derived from NB 2-4, contrary to the findings of Schmidt et al. (1997). It is medium-sized (6.4 um; n=4), dorsal, and migrates medially in abdominal segments. It projects contralaterally out the posterior root of the ISN before joining the SNa and forming an unusually robust "ribbon-like" synapse along the side of muscle 8 (Fig. 2-4). This pattern of innervation is a novel finding in Drosophila. However, a thin nerve branch, containing projections from the LBD and TA sensory cells, has been observed to connect SNa to ISN in third instar larvae (Haig Keshishian, personal communication). It appears that the motoneuron derived from NB 2-4 extends to muscle 8 by traveling in the same pathway and may correspond to the leukokinin cell identified in Calliphora by Cantera and Nassel (1992).
B. Interneurons:
All of the interneurons derived from this clone are local interneurons; we find approximately 8 interneurons per clone and find that they fall into two size populations: 50% of them were 3.1 microns in diameter (n=8 cells measured in 3 embryos) and the remaining 50% were approximately 4.8 microns in diameter (=7 cells measured in 4 embryos). We find a significantly different pattern of neurite projection for these cells than that reported by Schmidt et al, 1997. They report a single small interneuronal projection that extends posteriorly in the ipsilateral longitudinal connective. We find a robust projection extending across the midline to the contralateral longitudinal connective in the anterior commissure. In 7/8 clones, the motoneuron appeared to be fasciculated with these interneuronal projections until the midline. On reaching the contralateral longitudinal connective, the interneuronal projection turns anteriorly and extends to the anterior segment border. This is identical to the NB 2-4 interneuronal projections determined by eagle-kinesin-lacZ staining (Higashijima et al, 1996).
One interneuron migrates medially away from the clone, similar to the motoneuron. It appears to be a bipolar interneuron, previously unheard of in insect systems (Burrows, 1996). It can have one projection extending a short distance anteriorly (11 microns, n=2) and a second more robust projection extending posteriorly (32 microns, n=3) in the ipsilateral longitudinal connective (Fig 2-4 B,C(arrow)). The posterior projection frequently formed a loop at its terminus (Fig 2-4C,D).
References:
Bopp, D., Jamet, E., Baumgartner, S., Burri, M., and Noll, M. (1989). Isolation of two tissue-specific Drosophila paired box genes, Pox meso and Pox neuro. EMBO J 8(11):3447-57.
Broadus, J., Skeath, J.B., Spana, E. P., Bossing, T., Technau, G.M., and Doe, C.Q. (1995). New neuroblast markers and the origin of the aCC/pCC neurons in the Drosophila central nervous system. Mech Dev 53: 393-402.
Buescher, M., and Chia, W. (1997). Mutations in lottchen cause cell fate transformations in both neuroblast and glioblast lineages in the Drosophila embryonic central nervous system. Development 124 (3). 673-81.
Burrows, M. (1996). Oxford University Press, The Neurobiology of an Insect Brain. New York
Cantera, R., and Nassel, D. R. (1992). Segmental peptidergic innervation of abdominal targets in larval and adult dipteran insects revealed with an antiserum against leucokinin I. Cell Tissue Res 269: 459-471.
Chu-LaGraff, Q., Schmid, A., Leidel, J., Broenner, G., Jaeckle, H., and Doe, C. Q. (1995). huckebein specifies aspects of CNS precursor identity required for motoneuron axon pathfinding. Neuron 15: 1041-1051.
Cui, X., and Doe, C.Q. (1992). ming is expressed in neuroblast sublineages and regulates gene expression in the Drosophila central nervous system. Development 116(4): 943-52.
Cui, X., and Doe, C.Q. (1995). The role of the cell cycle and cytokinesis in regulating neuroblast sublineage gene expression in the Drosophila CNS. Development 121(10): 3233-43
Doe, C. Q. (1992) Molecular markers for identified neuroblasts and ganglion mother cells in the Drosophila central nervous system. Development 116: 855-863.
Higashijima, S-i., Shishido, E., Matsuzaki, M., and Saigo, K. (1996). eagle, a member of the steroid receptor gene superfamily is expressed in a subset of neuroblasts and regulates the fate of their putative progeny in the Drosophila melanogaster CNS. Development 122: 527-36.
Isshiki, T., Takeichi, M., and Nose, A. (1997). The role of the msh homeobox gene during Drosophila neurogenesis: implication for the dorsoventral specification of the neurectoderm. Development 124(16): 3099-3109.
McNeill, H., Yang, C.H., Brodsky, M., Ungos, J., and Simon, M.A. (1997). Mirror encodes a novel PBX-class ofhomeoprotein that functions in the definition of the dorsal-ventral border in the Drosophila eye. Genes Dev 11(8): 1073-82.
Schmidt, H., Rickert, C., Bossing, T., Vef, O., Urban, J., and Technau, G. M. (1997). The embryonic Central Nervous System lineages of Drosophila melanogaster II. Neuroblast lineages derived from the dorsal part of the neurectoderm. Dev Biol 189: 186-204.
Yang, X., Bahri, S., Klein, T., and Chia, W. (1997). Klumpfuss, a putative Drosophila zinc finger transcription factor, acts to differentiate between the identities of two secondary precursor cells within one neuroblast lineage. Genes Dev 11(11):1396-1408.